
«Динозавровый горизонт Средней Азии и некоторые вопросы стратиграфии»
Изв. АН СССР. — 1944. — Сер. геол. — № 3. — С. 40—58.
Автор анализирует условия захоронения в среднеазиатских местонахождениях динозавровой фауны, их распространение и связь с красно-цветными континентальными свитами мезокайнозоя и приходит к заключению о вторичном залегании остатков динозавров во всех известных местонахождениях, обусловленном крупным процессом размыва и переотложения верхнемеловых свит, возраст которого предположительно определяется как эоценовый. Наличие этого процесса размыва определяет отсутствие континентального эоцена в областях кайнозойской аккумуляции осадков.
Остатки динозавров в Средней Азии стали известны только за последнее тридцатилетие. Развитие геолого-съемочных работ в период 1924—1934 гг. и более ясное представление о характере отложений показали наличие очень крупных местонахождений динозавров, разбросанных на весьма обширной площади, главным образом, в Приташкентском районе, южной окраине Голодной степи и Илийской долине. Раскопки, предпринятые Геологическим комитетом и Геологическим музеем Академии Наук ССОР, доставили большое число костей различных представителей динозавров. Однако, несмотря на разнообразие собранного материала, составить представление о фауне среднеазиатских динозавров оказалось невозможным. (Сильно поврежденные кости, разрозненные и беспорядочно смешанные внутри пласта, могли дать почву лишь для самых общих и поверхностных заключений.
В 1929 г., по предложению Академии Наук СССР, я осмотрел ряд местонахождений динозавров в Казахстане и Киргизии и пришел к выводу о вторичном залегании остатков в этих местонахождениях (3). В другой статье я отметил огромный масштаб накопления и захоронения остатков динозавров в Средней Азии (4).
Трактовка динозавровых местонахождений как находящихся во вторичном залегании была принята частью исследователей. Однако другие среднеазиатские геологи, основываясь на сравнительно хорошей сохранности костей динозавров в приташкентских местонахождениях (Сары-Агач, Джилга), продолжали считать эти местонахождения коренными и их фауну датирующей возраст слоев. Благодаря постоянству и широкому распространению горизонта с костями динозавров в Приташкентском районе, этот горизонт является маркирующим и выделен в 1926 г. под названием динозаврового.
С местонахождениями Приташкентского района сходно местонахождение Биссекты в пустыне Кызыл-кум, открытое А.Ф. Соседко и являющееся крайней западной тачкой нахождения остатков динозавров из известных в настоящее время.
Наибольшее количество остатков было доставлено из местонахождений Илийской долины и Приташкентского района, а также и из Биссекты. Общее число костей, хранящихся в Палеонтологическом музее Академии Наук СССР и Центральном геологическом музее им акад. Чернышева в Ленинграде, более 1 500 номеров.
Несмотря на такой большой объем сборов, фауна из динозавровых местонахождений Средней Азии известна преимущественно по предварительным, зачастую весьма провизорным, определениям А.Н. Рябинина, А.П. Гартман-Вейнберг и моим.
Детальная монографическая обработка материалов до сих пор не произведена по причинам плохой сохранности материала. Исключение составляет работа А.Н. Рябинина (6), который пытался описать часть сборов динозавров из Приташкентского района (ст. Сары-Агач). В этой работе А.Н. Рябинин определил и описал остатки динозавров группы Ornithischia, не коснувшись хищных динозавров и зауропод. Наиболее определимыми он считает остатки траходонтных динозавров семейства Trachodontidae (Hadrosauridae) и подсемейства Lambeosaurinae, которые относятся к двум родам. Один род — совершенно новый — Jaxartosaurus aralensis, второй является новым видом; монгольского Bactrosaurus из слоев Ирен-Дабасу. Остатки стегозавров и, возможно, дератопсид только весьма поверхностно отмечаются автором, как очень редкие и неполные, а потому и неопределимые сколько-нибудь точно. Таким образом, из всего обширного материала, собранного в приташкентских местонахождениях, в монографии А.Н. Рябинина определены только остатки шлемоносных траходонтов — небольшой по объему группы ламбеозавров.
Однако, если подвергнуть критическому пересмотру фактический материал, легший в основу описания А.Н. Рябинина, то точность и этих определений придется считать мало yдoвлeтвоpитeльнoй. В самом деле, разрозненные остатки скелета Jaxartosaurus — нижняя челюсть, позвонки, таз, плечо и берцовая кость, — по мнению самого автора, лишь предположительно относятся к устанавливаемому им роду. Для всех упомянутых элементов скелета А.Н. Рябинин не дает и не может дать ни одного характерного признака, несомненно отличающего или несомненно сближающего их с ранее известными формами. Обломок черепа, послуживший А.Н. Рябинину основой для описания нового рода, не может выявить черт строения, определенно отличающих его от таких форм, как Hypacrosaurus, Bactrosaurus и даже Cheneosaurus. Части верхней и нижней челюстей траходонтного динозавра, описанные А.Н. Рябининым как новый вид Bactrosaurus, еще более неясны в своей систематической принадлежности. Единственным критерием для отнесения остатков к роду Bactrosaurus А.Н. Рябинину послужило число зубных бороздок на dentale. Как известно, этот признак у траходонтов весьма неустойчив и подвержен индивидуальным и, особенно, возрастным вариациям. Поэтому отнесение фрагмента к роду Bactrosaurus нужно считать мало обоснованным. Таким образом! рассмотрев работу А.Н. Рябинина — единственную монографию, посвященную динозаврам Средней Азии, приходим к выводу; что динозавровая фауна Приташкентского района еще не дала ни одной определимой формы. Систематическое положение, географические связи и эволюционная ступень среднеазиатских динозавров еще совершенно темны.
Новый род Jaxartosaurus, выделенный Рябининым, не говоря уже о видах, недостаточно обоснован и не может служить опорным при анализе фаун других местонахождений. Разумеется, совсем не исключена возможность, что в фауне динозавров Средней Азии мы можем встретить новые формы, знаменующие обособления широко распространенных групп. Однако выделение новых форм должно быть основано на гораздо более полном материале и только в этом случае будет убедительно.
Малые результаты проделанного А.Н. Рябининым исследования не случайны и определяются отмеченным выше плохим состоянием и разрозненностью остатков, характерными для всех среднеазиатских местонахождений без исключения. Находящийся в моем распоряжении материал раскопок Академии Наук СССР и двукратных сборов из Кызыл-кумов равным образом дает возможность сделать лишь самые общие определения крупных систематических единиц, более мелких, чем семейства. Поскольку более полные материалы по динозаврам в Советском Союзе совершенно отсутствуют, то остается только одна возможность удовлетворительного определения обломочного материала среднеазиатских местонахождений — путем прямого сравнения костей с обширными коллекциями американских и других зарубежных музеев. По условиям военного времени эта возможность естественно отпадает.
Приведенный краткий обзор изученности динозавровой фауны Средней Азии заставляет признать стратиграфическое значение остатков динозавров условным. Несмотря на широкую распространенность, очень характерный облик и, следовательно, легкую полевую определимость, остатки динозавров могут служить лишь для идентификации отложений в разных районах. Вопрос о подлинном возрасте фауны динозавров приходится считать открытым. Последнее имеет совершенно особенное значение при явственно вторичном залегании костей динозавров во всех без исключения местонахождениях, известных в настоящее время.
Признавая невозможность точной датировки вмещающих отложений с помощью остатков динозавров, мы не можем пройти мимо огромного масштаба явления. Большие площади, занимаемые костеносными слоями, их распространение в самых различных местах Средней Азия и наконец чудовищное количество захороненных остатков, — все это указывает на то, что накопление остатков динозавров явилось крупным процессом, имеющим самостоятельное значение для геологической истории Средней Азии. Анализируя этот процесс с точки зрения тафономии, т. е. закономерностей захоронения животных остатков, мы можем получить некоторые данные для суждения о стратиграфическом значении динозавровых местонахождений и относящихся сюда общих вопросов геологической истории Средней Азии. К рассмотрению этих данных мы и переходим.
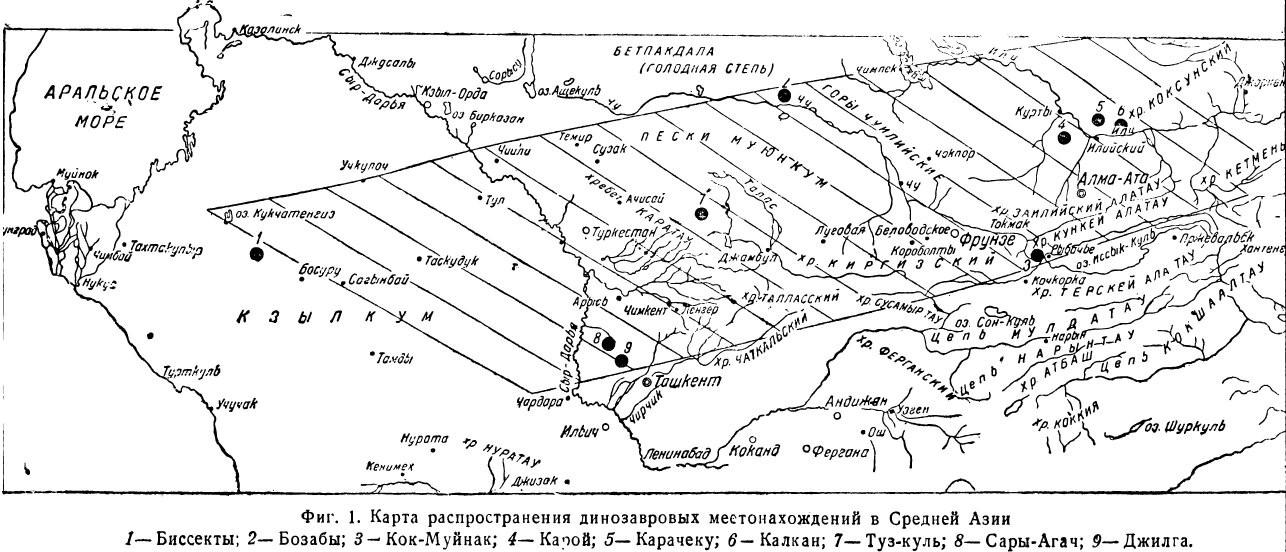
На территории Средней Азии в настоящее время известно девять крупных местонахождений (фиг. 1), в самом общем представлении располагающихся широтно вдоль северной окраины горных цепей. Ни одно из этих местонахождений не исследовано сколько-нибудь детально, большинство не имеет даже ясной геологической характеристики. Подвергавшиеся раскопкам местонахождения Карачеку, Калкан, Сары-Агач и Джилга (5, 6, 8, 9) не были исследованы специалистами, и мы не получили по ним данных, определяющих важнейшие тафономические элементы местонахождения. Изучение динозавровых местонахождений с точки зрения условий их образования. — большая и трудоемкая работа, составляющая задачу будущего. Краткая сводка достоверного фактического материала по девяти динозавровым местонахождениям, показанным на нашей карте, сводится к следующему (фиг. 1).
1. Местонахождение Биссекты (пустыня Кызыл-кум) у колодца Биссекты, к северу и западу от гор Букан-тау, в 100 км к юго-западу от оз. Кукча-Тенгиз. Обширная площадь, занятая размытыми и выходящими непосредственно на поверхность, горизонтально лежащими слоями песчаников и конгломератов с костями динозавров. Площадь костеносных слоев не менее 20 кв. км. Кости разрознены и беспорядочно перемешаны в пласту, частью окатаны и, в большинстве случаев, разломаны. Кости очень твердые, хорошей сохранности, с гладкой поверхностью, нередко вторично окремненные, часто с примесью закиси железа. Непосредственно кроющие свиты не наблюдались, мощность костеносного слоя неизвестна. По-видимому, костеносные песчаники залегают на размытой поверхности дислоцированных древних свит.
В составе фауны, по определению И.А. Ефремова, преобладают травоядные динозавры, группы Ornithischia семейства Trachodontidae (траходонты). Хищные динозавры (Saurischia, Carnosauria) также часто встречаются в фауне Биссектов и представлены как крупными хищниками типа Deinodontidae, так и мелкими Ornithomimidae. Имеются очень редкие и трудно определимые остатки зауропод и анкилозавров.
Кроме динозавров, много остатков черепах Cryptodira (сем. Trionychidae и семейства Testudinidae) и зубов акул, а также обломков окремнелой древесины хвойных деревьев.
2. Местонахождение Бозабы на южной окраине Голодной степи — правый берег- р. Чу, против озера Кокый, лежащего на левом (берегу Чу, в 90 км от поселка Пуляевка. В этом местонахождении костеносные слои значительно размыты и выходят непосредственно на дневную поверхность в виде гряды окаменевших костей и гальки, тянущейся на десятки километров. Покрывающие слои неизвестны, как и мощность костеносного горизонта, достигающая, однако, 3—4 м. Самый косте-носный пласт — грубые конгломераты с линзами песчаников, местами с гипсом. Кости сильно окатаны, с поврежденной трещиноватой поверхностью. Состав фауны неизвестен, так как сборы не производились. Доставленные отдельные фрагменты принадлежат траходонтам и карнозаврам. Местонахождение выделяется среди всех остальных громадными размерами.
3. Местонахождение Кок-Муйнак в долине p. Чу, между поселками Рыбачье и Быстровка, в средней части Боомского ущелья, на левой (по течению) стороне урочища Кок-Муйнак. В пределах большого разреза красноцветных третичных отложений находится изолированный небольшой массив конгломератов с костями динозавров, террасовидно прислоненный к сильно дислоцированным и метаморфизованным палеозойским отложениям. В верхней части костеносного горизонта, достигающего 6 м мощности, проходит линия надвига, по которой динозавровые слои перекрываются неогеновыми конгломератами. Детальные соотношения с окружающими породами не выяснены. Кости плохой сохранности, преимущественно в виде мелких обломков с трещиноватой, импрегнированной гипсом поверхностью. В числе представителей фауны могут быть отмечены, по определению И.А. Ефремова, траходонты, дейнодонты, орнитомимиды и черепахи семейства триониксов. Местонахождение Кок-Муйнак является самым небольшим и наиболее изолированным из всех известных. Костеносные слои его представляют собой островок, зажатый среди больших массивов палеозойских и третичных пород путем вертикальных тектонических нарушений.
4. Местонахождение Карой в урочище Карой, у колодца Сары-кудук около Аусу-кудук, в пределах левобережья р. Или, на северо-запад от ст. Илийск (Турксибская железная дорога). Террасовидно прислоненный к массиву палеозойских пород островок размытых красновато-желтых конгломератов с костями динозавров. Кроющие слои отсутствуют. Мощность свыше 3 м, но точно не выяснена. Кости светлосерого цвета; очень сильно разломаны, частично окатаны, с разрушенной трещиноватой поверхностью. В цементе костеносной породы — желтый песок, сцементированный гипсом. Гипсом проникнуты и все трещины костей. Состав фауны полностью неизвестен, однако несомненно присутствие траходонтов и орнитомимид, а также триониксов из черепах.
5. Местонахождение Карачеку в области левобережья р. Или — сухой широкой долины, к юго-востоку от бывшей почтовой станции Карачеку. Горизонтально лежащие слои черных конгломератов с галькой разной крупности, с резко выраженной косой слоистостью, вскрывающиеся непосредственно на дневной поверхности. В глубоких промоинах и ложках видно, что конгломераты с костями динозавров мощностью около 5 м залегают на размытой поверхности метаморфизованных сланцев палеозоя. Общая площадь костеносного поля не менее 4 кв. км состоит из однообразных конгломератов, террасовидно прислоненных к древним породам. Галька в конгломерате из подстилающих метаморфических сланцев, цемент из песка с закисью железа и марганца. Кости большей частью крупных размеров, всегда поврежденные и разрозненные, с разрушенной поверхностью, частью окатанные, пропитанные закисью железа и марганцовыми соединениями. Встречаются обломки древесных стволов, также пропитанные железом и марганцем.
В составе фауны, по определению А.П. Гартман-Вейнберг и И.А. Ефремова, — Tpachodontidae (траходонты), Deinodontidae, Ornithomimidae, а также сравнительно большое количество остатков зауропод? (Titanosauridae), стегозавров? (Stegosauria) и анкилозавров (Acanthopholldae). Имеются многочисленные обломки щитков черепах семейства Trionychidae.
6. Местонахождение Калкан в пределах правого коренного берeгa р. Или, у подножья юго-западной стороны горной группы Кши-Калкан, близ родников Калкан.
Костеносный горизонт частью выходит на дневную поверхность, частью прикрыт желтыми косослоистыми песками с прослоями мелкой гальки видимой мощности около 5 м. Самый костеносный пласт — рыхлый грубый конгломерат с крупной галькой кристаллических и метаморфических пород, цементированных гипсоносным желтым песком. Конгломерат залегает на головах размытых палеозойских отложений и в средней восточной части местонахождения прислоняется непосредственно к палеозойскому массиву Кши-Калкан. Залегание горизонтальное, полная мощность костеносного конгломерата неизвестна, но, вероятно, щ менее 3—4 м. Площадь костеносного пласта около 6 кв. км. Кости светлосерого цвета, разломанные и беспорядочно перемешанные с галькой. Поверхность костей разрушена и трещиновата, частью покрыта гипсовыми натеками, реже — железистой корочкой. Много окатанных обломков. Довольно часто встречаются обломки окремнелой древесины, также покрытые гипсовыми натеками. В составе фауны, по определению А.П. Гартман-Вейнберг и И.А. Ефремова, преобладают траходонтные динозавры Traidiodontidae. Много остатков хищных Carnosauria, вероятно, представителей семейства Deinodontidae, а также Ornithomimidae. Очень интересно отметить присутствие явственных представителей ортопод — семейства Ceratopsidae,? Stegosauridae и Acanthopholidae. В этом местонахождении встречены впервые среди всех остальных несомненные крупные цератопсиды типа Triceratops. Имеются большие спинные шипы, отличные от известных анкилозавров и крайне сходные с типичными стегозаврами. Найдены представители зауропод, вероятно, относящиеся к семейству Titanosauridae. Кроме динозавров, много обломков панцыря черепах, в подавляющем большинетве случаев Trionychidae, хотя имеются и Testudinidae.
7. Местонахождение Туз-куль в пределах восточных предгорий хребта Кара-тау, у озера Туз-куль на Тузкульском месторождении целестина. Желтая свита рыхлых песчаников и конгломератов с когтями динозавров залегает на размытой поверхности толщи красноцветных гипсоносных и целестиноносных глин каменноугольного возраста.
Местонахождение недавно открыто и еще не обследовано, поэтому основные данные отсутствуют. По характеру костеносных пород и типу фоссилизации костей динозавров местонахождение Туз-куль чрезвычайно сходно с местонахождением Калкан. Нужно думать, что основная характеристика этого местонахождения не отличается от только что приведенной для Калкана. Пока из Туз-куля известны только траходонтные динозавры семейства Trachodontidae, но нет оснований предполагать, что его фауна будет резко отличаться от всех остальных.
8 и 9. Группа местонахождений Приташкентского района (Сары-Агач и Джилга) имеет более полную геологическую характеристику. Здесь обнаружен ряд выходов костеносных пород в холмистой степи Чули. Горизонт с костями динозавров простирается на обширной площади, сложенной размытыми отложенями мела и палеогена. Наиболее богатыми остатками динозавров являются следующие пункты: район урочища Сары-Агач к югу и западу от железной дороги (ст. Ченгельды Ташкентской железной дороги, колодец Узун-кудук в хребте Алым-тау) район колодца Кырк-кудук (хребет Казы-гурт); колодец Суксун-кудук на Ширапханинской дороге к северо-западу от долины Сары-Джилга и др. В некоторых местах выходы костеносного горизонта тянутся на протяжении до 20 м (Алым-тау) в виде узкой полосы или же выступают на дневную поверхность широкими участками в 3—4 кв. км (урочище Тюльку-су).
Последние исследователи (5) помещают динозавровый горизонт в верхи чанакской свиты и относят к сеноману, а не к турону, как это было сделано Борнеманом (2). По их мнению, динозавровый горизонт согласно прикрывается меловыми отложениями дарбазинской свиты, мощностью свыше 100 м с морской фауной сенона. Динозавровый горизонт согласно подстилается конгломератами, песчаниками и глинами чанакской свиты около 400 м мощности, а чанакская свита в свою очередь согласно подстилается азатбашской красноцветной свитой нижнего мела небольшой мощности, залегающей непосредственно на размытой поверхности палеозойских пород. Однако на самом деле нигде в пределах выходов несомненно костеносного горизонта мы не наблюдаем непосредственного контакта со слоями, охарактеризованными фауной беспозвоночных. При очень запутанной картине сложных фациальных переходов континентальных и морских отложений Чулей, осложненной тектоникой и сильным размывом, точность приведенных сопоставлений является сомнительной. Несомненными являются фактически наблюдавшиеся в обнажениях разрезы, из которых (рассмотрим наиболее характерный, приведенный А.Н. Рябининым в монографии (Давлет-сай; 6).
1. Известняки с эоценовой (фауной, частично размытые.
2. Зеленые и красные глины, внизу переходящие в серые песчаники — 82 м.
3. Розовый песчаник — 3 м.
В другом месте выходят:
4. Красные глины — 12 м.
б. Светлозеленые известковистые глины — 0.5 м.
6. Серые песчаники, переполненные гастроподами — 12 м.
7. Красные косослоистые песчаники — 17 м.
8. Красные песчаники с прослоями красных глин, в низах содержащие стволы хвойных деревьев, сильно окатанные кости динозавров и поврежденные раковины гастропод и пелеципод — 90 м.
9. Красный брекчиевидный известняк с костями динозавров, черепах и рыб — 9 м.
В других участках костеносный горизонт, имея тот же характер сохранности и фоссилизации костей, состоит из песчаников, конгломератов и плотного розового («строительного») песчаника. Наиболее часто и в наибольших количествах кости встречаются в конгломератах.
Тип сохранности костей весьма сходен с описанным для местонахождения Биссекты. Кости плотные, большей частью с гладкой, хорошо сохранившейся поверхностью. Все кости в той или иной степени повреждены, разломаны и беспорядочно перемешаны внутри костеносного пласта. Много окатанных фрагментов. Многие кости несут явственные признаки вторичного окремнения.
Состав фауны неоднократно указывался А.Н. Рябининым, причем определения в отдельных случаях давались до вида. Выше, в разборе работы А.Н. Рябинина, мы показали слабую обоснованность детальных определений этого исследователя. Поэтому приведем список фауны по его данным, исключив сомнительные, указанные самим автором под вопросом, видовые и родовые определения.
В таком более достоверном освещении фауна динозаврового горизонта Приташкентского района имеет следующий состав: преобладают остатки траходонтных динозавров семейства Hadrosauridae, (Trachodontidae) и подсемейства Lambeosaurinae; также довольно часты остатки карнозавров семейств Deinodontidae и Ornithomimidae; очень редки остатки зауропод, стегозавров и цератопсид, которые могут быть отнесены к семейству Titanosauridae, Nodosauridae и Ceratopsia sensu lato; много остатков черепах, преобладают Cryptodira (сем. Trionychidae и Dermatemydidae); имеются сомнительные остатки черепах, отнесенные А.Н. Рябининым к Ampnichelydia; многочисленны позвонки крупных костистых рыб (? Portheus).
Некоторые участки костеносного горизонта отличаются большим количеством окаменевших стволов хвойных. В других участках отмечается много раковин гастропод и пелаципод, среди последних толстостенные тригонии.
Все рассмотренные нами девять местонахождений можно отнести к двум основным типам, выделяющимся достаточно отчетливо. Первый тип, к которому принадлежит местонахождение Биссекты и приташкентские местонахождения, отличается следующими чертами.
1. Сравнительно хорошая сохранность костного материала. Кости с гладкой поверхностью, большей частью только разломаны, но не окатаны. Процент сильно окатанных костей невелик.
2. Костеносные слои представлены не только конгломератами, но и песчаниками и известняками.
3. Костеносные слои залегают внутри довольно мощной свиты генетически связанных с костеносным пластом пород, содержащих также редкие обломки сильно окатанных костей и крупные стволы деревьев достигающие нескольких метров длины.
4. В составе фауны местонахождений характерно большое количество остатков рыб (зубы акул, костистые рыбы) и крайняя редкость остатков динозавров, цератопсид и зауропод.
5. Имеется несомненная, хотя и еще далеко неясная, связь костеносных пород с окружающими свитами.
Второй тип, характерный для всех восточных местонахождений среднеазиатских динозавров, включает в себя местонахождения Бозабы, Кок-Муйнак, Карой, Карачеку, Калкан и Туз-куль. Нужно подчеркнуть, что все эти местонахождения сходны между собой по геологическим условиям залегания и, особенно, по характеру сохранности и фоссилизации костного материала. Обогащение железом и марганцем, отмеченное для местонахождений Карачеку, нисколько не меняет всей остальной картины поразительного единства второго типа.
Существенными чертами второго типа являются:
1. Залегание в виде отдельных, уцелевших от размыва массивов, непосредственно на палеозойских, сильно дислоцированных породах.
2. Довольно сильная степень разрушения костного материала: поверхность костей потерта, изъедена выветриванием и трещиновата; большой процент окатанных фрагментов.
3. Костеносный слой является, за редкими исключениями, грубым конгломератом с крупными гальками, сравнительно слабо цементированным.
4. В составе фауны отсутствуют заметные остатки рыб. Значительно более часты, чем в местонахождениях первого типа, находки стегозавров, цератопсид и зауропод.
5. Не имеется отчетливой связи с окружающими красноцветными породами.
Оба рассмотренных типа местонахождений динозавров являются вторичными. Вторичное значение костного материала безусловно доказывается состоянием и характером фоссилизации костей. Кости несут сле-ды выветривания, разломы и потертости, несомненно происшедшие после фоссилизации, т. е. тогда, когда кости уже были окаменелыми и процесс диагенеза и литификации костеносной породы был полностью закончен. Нанести столь сильные повреждения громадным костям динозавров могли только весьма сильные потоки и то лишь при обязательном условии хрупкости костей, т. е. опять-таки при законченной фоссилизации костного вещества. Другие признаки — беспорядочное перемешивание костей самых различных животных, отсутствие естественной для коренных местонахождений биологической и гидромеханической сортировки остатков, абсолютное отсутствие цельных элементов скелета, повреждения поверхности костей с несомненностью подтверждают перемыв и деструкцию коренных местонахождений.
Местонахождения первого типа — приташкентские и Биссекты — образовались за счет перемыва коренных местонахождений, находившихся недалеко от места отложения перемытого материала. Сохранность поверхности костей в этих местонахождениях свидетельствует о том, что перемыв происходил в условиях большого количества воды, покрывавшей мощным слоем перемываемые участки, которые не вскрывались на субъаэральной поверхности. Процесс перемыва был безусловно длительным и происходил при неуклонном убывании силы потоков, закончившись в отложении красных косослоистых песков дельтового типа. Эти последние отложения содержат не только сильно окатанные мелкие фрагменты костей, но также и большие древесные стволы, перенесенные в плавающем состоянии и фоссилизированные безусловно позднее, чем кости динозавров. Кости рыб Portheus и зубы акул подтверждают приведенную трактовку генезиса местонахождений первого типа.
Таким образом, мы приходим к выводу, что местонахождения динозавров первого типа образовались в непосредственной близости большого водного бассейна, вероятно моря. Давно образовавшиеся коренные местонахождения динозавров находились недалеко и были нацело перемыты, очевидно, при перемещении дельты крупного потока.
Генезис местонахождений второго типа существенно отличается. Коренные местонахождения динозавровой фауны были после фоссилизации выведены на субъаэральную поверхность и подвергались выветриванию и размыванию, в результате чего все вмещающие породы были нацело уничтожены и на месте остались лишь кости динозавров, как наиболее крупная тяжелая фракция. Впоследствии кости динозавров перемещались и отлагались потоками большой силы, осаждавшими вместе с тем и крупную гальку. Кости динозавров в образовавшихся отложениях пролювиального типа эквивалентны гальке вмещающих конгломератов и являются, как отмечено, остаточной фракцией от нацело уничтоженных древних отложений, содержавших коренные местонахождения. При этом представлении вполне закономерно залегание местонахождений второго типа в виде изолированных островков или массивов. Различие в составе фауны обоих типов динозавровых местонахождений обязано, конечно, первичному составу фауны в коренном залегании и объясняется разностью фациальных условий образования коренных местонахождений. Интересно отметить, что наличие стегозавров и цератопсид в местонахождении второго типа может обозначать их коренное образование в более удаленных от морского берега и возвышенных областях, что согласуется и с континентальным характером последующих процессов их перемывания.
Фауна всех динозавровых местонахождений обоих типов имеет отчетливый верхнемеловой возраст. Единственным признаком более древних форм являются типичные для стегозавров спинные пластины. Настоящие стегозавры были юрскими формами и окончательно исчезли в нижнем мелу, однако, на основании столь неполных остатков нельзя сделать никаких достоверных выводов. Более вероятно будет предположить, что в среднеазиатской фауне динозавров могли быть или новые формы стегозавров более поздней эволюционной стадии, или же эти остатки принадлежат новым, еще неизвестным, формам анкилозавров — группе, сменившей стегозавров в меловом периоде. Крупные цератопсиды и карнозавры, а также типичные орнитомимиды характерны для наиболее высоких горизонтов верхнего мела — ларамийской формации или датского яруса. Шлемоносные траходонты, дейнодонты, титанозавриды являются характерными формами несколько более низких слоев — приблизительно верхов сенона. Остатки черепах менее характерны и датируют время от верхнего мела до олигоцена.
До того времени, когда мы путем более полных находок и детальных исследований сможем оценить эволюционную стадию различных форм динозавровой фауны Средней Азии, нужно считать, что эта фауна имеет возраст сенон-датский ярус (ларами), в среднем — верхний сенон. Этот возраст, разумеется, может относиться только к коренным местонахождениям динозавровой фауны, ныне исчезнувшим. Для перемытых вторичных местонахождений возраст может оказаться, очевидно, любым и непременно более поздним. Априорно можно сказать, что для местонахождений первой группы их возраст будет близок во времени к возрасту коренных. Для местонахождений второй группы, по-видимому, между образованием коренных и отложением вторичных местонахождений прошел более значительный промежуток времени. Это предположение основано лишь на косвенных указаниях.
Суммируя фактический материал по динозавровым местонахождениям Средней Азии, мы должны подчеркнуть, что не только коренные местонахождения, но и самые толщи, могущие быть вместилищем таковых, в пределах Средней Азии неизвестны. В наиболее «близких» к коренным местонахождениям первой группы подстилающие слои достигают довольно большой мощности (400 м), но нигде не содержат остатков позвоночных. Для второй группы отсутствие возможностей нахождения поблизости коренных скоплений костей еще более убедительно.
Одно из основных положений тафономии гласит, что в геологической летописи сохраняются лишь производные крупных процессов. Мелкие по размерам и длительности существования фации стираются во времени неизбежными процессами деструкции.
Выше неоднократно отмечался гигантский масштаб накопления остатков динозавров, запасы которых в местонахождениях исчисляются сотнями тысяч или даже миллионами тонн. Для образования таких больших вторичных местонахождений нужны огромные коренные скопления костей, в которых захоронялось несметное количество животных. Само собой разумеется, что подобные скопления животных остатков могли образоваться лишь в случае обилия фауны и достаточной длительности благоприятных для захоронения условий. Другими словами, в эпоху обитания среднеазиатских динозавров в верхнем мелу должны были существовать длительные в смысле времени и широко развитые в смысле пространства условия расцвета и захоронения фауны. Эти фациальные условия верхнего мела не могли не отразиться в геологической летописи и до нас должны были дойти мощные, преимущественно дельтовые, отложения с крупными местонахождениями фауны и флоры. На самом деле, в пределах Средней Азии при довольно хорошей геологической изученности мы не встречаем ничего подобного. Нужно думать, что на крупный процесс образования пояса дельтовых осадков с коренными местонахождениями верхнемеловой фауны во времени наложился другой, не менее крупный, процесс — размыв указанных осадков и образование остаточных вторичных скоплений перемытых костей. Только благодаря огромным размерам первичных местонахождений остатки динозавров дошли до нас в большом количестве, хотя и подвергшиеся деструкции. С другой Стороны, именно широкая распространенность, большой масштаб процесса размыва обеспечили сохранение во времени его продуктов — осадков с перемытыми костями динозавров. Действительно, крайнее сходство всех местонахождений, особенно второго типа, между собой ярко доказывает общность условий их образования, а распределение местонахождений на огромной площади (фиг. 1) показывает пространство, захваченное процессом перемыва верхнемеловых костеносных свит.
Определение возраста процесса перемыва устанавливает и истинный возраст динозавровых местонахождений.
Вне всякого сомнения, пристальное изучение геологической истории Средней Азии может дать материал для анализа геологического времени такого, крупного процесса. В эту сторону и должны быть направлены стратиграфические исследования мезозойских красноцветных толщ. Мы, со своей стороны, попытаемся наметить некоторые перспективы определения возраста динозавровых местонахождений.
Для суждения об истории соответствующих областей Средней Азии в эпоху образования коренных динозавровых местонахождений пока имеется очень небольшое количество данных. Динозавровые слои залегают на размытой поверхности палеозоя, или же, как в местонахождениях первой группы, они отделены от поверхности размыва сравнительно незначительной толщей однообразных мертвых пород.
В превосходной, но еще не опубликованной диссертации С.С. Шульца дан общий обзор условий седиментации в позднемезозойское и третичное время для интересующей нас области. Автор приходит к выводу, что красноцветные континентальные толщи Тянь-Шаня в подавляющем большинстве случаев являются третичными образованиями. Сопоставляя детальный анализ новейшей тектоники с показаниями фауны и флоры, С.С. Шульц убедительно доказывает, что главнейшие поднятия хребтов Тянь-Шаня в общем начались в верхах палеогена или низах неогена. С этими поднятиями связана аккумуляция мощных третичных континентальных отложений в крупных межгорных впадинах. Возраст базальных свит третичных толщ Тянь-Шаня нужно считать или верхнеолигоценовым или миоценовым. Наибольшей, нередко километровой, мощности достигают неогеновые отложения. В большинстве случаев третичные отложения ложатся на поверхность древней денудации, на палеозойские породы. В одной из наиболее крупных депрессий — Иссык-кульской котловины — сохранились частично юрские угленосные отложения, находившиеся в тесной связи с угленосной юрой Кульджи через чрезвычайно сходные отложения в долине Текеса и Кетменском хребте. Курские отложения совершенно не содержат грубообломочных конгломератовых фаций, столь характерных для третичных отложений, и входят в состав отложений, слагающих поверхность древней денудации. На размытой поверхности юры залегают красноцветные третичные осадки.
К западу, в еще более крупной Ферганской котловине, известны меловые отложения. Нижняя красноцветная свита до 1000 м мощности (сильно изменчивой), по-видимому, континентального происхождения, сложена яркоокрашенными конгломератами и песчаниками, иногда с базальтовыми покровами. Верхняя свита морская, фаунистически охарактеризованная (турон-сенон), сложена глинами, известняками и песчаниками до 500 м мощности. В северо-восточном направлении морские осадки верхнего мела сменяются континентальными. Морские палеогеновые отложения ложатся без видимого углового несогласия на морской мел или, имея более широкое распространение, на нижележащие породы.
К востоку от Ферганской котловины морской палеоген не известен ни в хребтах, ни во впадинах Тянь-Шаня.
Палеогеновое море в своем распространении не было связано с современным положением главных хребтов и впадин Тянь-Шаня.
Еще в юрское время Тянь-Шань был областью сноса. Тектонические движения в мезозое и палеогене не были интенсивными и почти полностью компенсировались размывом и аккумуляцией, не нарушая общего равнинного характера страны.
Сравним приведенные из работы С.С. Шульца палеогеографические данные и наши сопоставления с наиболее богатыми фауной позвоночных разрезами континентальных толщ мезокайнозоя Монголии. Как известно, значение Монголии для эволюции наземных позвоночных в эпоху верхнего мезозоя и кайнозоя было весьма велико. Монголия была одним из центров развития наземных позвоночных в этот период, так как центральноазиатское плато было континентом с юрского периода до наших дней. Поэтому в отложениях здесь представлены континентальные фации, сохранившие остатки фауны мела и третичного периода, до сих пор известной только из береговых отложений Европы и Америки. На размытую поверхность древнего юрского континента Монголии («великое несогласие», по терминологии американских исследователей) ложатся континентальные отложения очень низких горизонтов нижнего мела. Эти отложения, представленные тремя свитами (снизу вверх) — Ондай Сайр, Он-Гонг и Оши, общей мощностью около 900 м, сложены песками, конгломератами, глинами и тонкими известняками со своеобразной фауной динозавров. Наиболее характерны крупные диплодокообразные зауроподы и предковые формы ортопод и дейнодонтов (Protiguanodon, Psittacosaurus, Prodeinodon, Asiatosaurus). Выше согласно залегают еще восемь свит нижнего мела, общей мощностью около 1200 м с разнообразной фауной динозавров (зауроподы, игуанодонты), крокодилов, черепах и пресноводных беспозвоночных. На нижнемеловые отложения, без видимого несогласия, налегают две верхнемеловые свиты: Ирен-Дабасу и Джадохта, общей мощностью около 250 м. В этих свитах, сложенных красными песками, красными и серыми глинами и песчаниками, заключена богатейшая фауна динозавров — игуанодонтов, дейнодонтов, орнитомимид, своеобразных цератопсид — Protoceratops, а также меловых млекопитающих. Эта фауна, более примитивная, чем фауна самых верхов мела в Европе и США, относится исследователями ориентировочно к сенону. На верхний мел Монголии налегают с слабым несогласием (смещения заметны близ хребтов Монгольского Алтая) красные и бурые глины, серые грубые пески палеоцена свиты Гашато около 100 м мощности, с фауной млекопитающих Palaeostylops, весьма сходной с соответствующей фауной США. На палеоцен согласно налегают шесть свит эоцена, общей мощностью около 300 м, красных глин, белых и серых песков и песчаников с замечательной фауной титанотериев и халикотериев. На этих отложениях согласно лежат весьма сходные породы олигоцена — семь свит мощностью 400 м с фауной титанотериев и гигантских носорогов — белуджитериев, затем небольшой мощности зеленые пески и глины миоцена (свита Лох, 30 м мощностью) с фауной Serridentinus. Выше идут еще свиты плиоцена около 700 м мощности и плейстоцена (около 100 м).
Проявления складчатости, постепенно затухающие, отмечаются почти во всех свитах мела. Базальтовые потоки характерны для нижнего мела, палеоцена и самых верхов олигоцена. В остальных свитах проявлений вулканизма не наблюдается.
Юрские отложения, входящие в состав древнего континента Монголии — поверхности великого несогласия — угленосны и очень сходны с континентальной юрой Забайкалья, Кульджи, Кетменя и вышеуказанными угленосными отложениями юры в Иссык-кульской котловине не только по характеру отложений, но и по возрасту (нижняя юра-лейас). По всей вероятности, предистория мелового периода на континентальном участке Средней Азии была весьма сходна с историей юрского континента Центральной Азии и, в первую очередь, Монголии. Подобно Монголии, юрские угленосные отложения Средней Азии входят в состав древней денудационной поверхности, на которую несогласно ложатся осадки всех последующих периодов аккумуляции. Нужно думать, что исследования весьма сходных соотношений на территории Синьцзяна покажут, что эта область, промежуточная между Монголией и нашей Средней Азией, имела такую же историю континентальных областей.
Совершенно несомненно распространение динозавровой фауны по всем областям Азиатского мезозойского континента. Огромные количества остатков, захороненные в наших среднеазиатских местонахождениях, знаменуют такое развитие и расцвет динозавров, какие отнюдь не могли иметь места на обособленном и сравнительно небольшом участке мезозойской суши, очерченном распространением среднеазиатских местонахождений. Отсюда ясно единство развития динозавровой фауны на обширном пространстве Азиатского материка, да и весьма многих смежных стран. Поэтому среднеазиатская динозавровая фауна является лишь позднейшим дериватом обширного центра развития мезозойской фауны на Азиатском материке, но дериватом, показывающим могучий расцвет динозавров с поражающей численностью индивидов.
Исходя из этих соображений, мы вправе предполагать известную общность основных физико-географических условий для мелового континента Центральной и Средней Азии, и следовательно, видеть в континентальных толщах Монголии эталон для восстановления в общих чертах стершегося в Средней Азии куска геологической летописи. Первым опорным пунктом является вышеотмеченное сходство юрских отложений и юрской денудационной поверхности, вторую точку опоры составляет богатство, разнообразие и общее сходство состава фауны динозавров. Как мы увидим в дальнейшем изложении, имеется еще один опорный пункт, связующий историю нашей Средней Азии с крупными этапами истории мезокайнозойского Азиатского материка.
При прямом сравнении континентальных отложений Монголии и рассматриваемой области Средней Азии, аккумулированных над общей юрской денудационной поверхностью, сразу же бросается в глаза выпадение фаунистически охарактеризованного континентального мела и эоцена. Отложение континентальных свит в Средней Азии, в областях распространения динозавровых местонахождений (главным образом второго типа), начинается с олигоцена. Выше следует мощная серия континентальных пород до плейстоцена включительно. Таким образом, от верхнего мела и всего эоцена на участках древней мезозойской суши Средней Азии уцелели только перемытые остатки громадных местонахождений. Поскольку коренные местонахождения отлагались в самых верхах мела, естественным выходом из всего изложенного будет отнесение процесса крупного размыва к эоценовому времени. В этом случае отсутствие мощного, фаунистически охарактеризованного континентального эоцена в Средней Азии и вообще во всех смежных районах будет вполне закономерным. Нужно думать, что грубые динозавровые конгломераты и являются единственно известными континентальными отложениями эоцена, удержавшимися во время крупной эоценовой денудации на обширной области сноса.
Если мы обратимся к огромной площади развития третичных отложений Казахстана, непосредственно примыкающей с севера к полосе развития динозавровых конгломератов, то мы увидим много весьма важных и интересных соотношений. Наряду с очень богатыми местонахождениями третичных млекопитающих, распространенными почти во всех районах развития третичных отложений и охватывающими период от среднего олигоцена до позднего миоцена и плиоцена, ярко выделяется отсутствие эоценовых отложений и эоценовой фауны млекопитающих. И здесь континентальная история геологических образований начинается с олигоцена, с полным выпадением всего эоцена. При этом еще более интересно, что нижние свиты континентальных третичных отложений Казахстана содержат очень характерную фауну гигантских носорогов — белуджитериев, — так называемые индрикотериевые слои. Эта фауна очень типична для олигоцена Монголии и может служить третьим опорным пунктом, связующим и более поздние этапы геологической истории Азиатского материка в Монголии и нашей Средней Азии.
В новейшей сводке акад. А.А. Борисяка (1) подробно изложены сравнительные данные по генезису казахстанских и монгольских местонахождений третичных млекопитающих. А.А. Борисяк приходит к выводу, что возраст индрикотериевых слоев Казахстана, ранее принимавшийся за верхнеолигоценовый, должен быть понижен до среднего олигоцена. Таким образом, известная нам континентальная история кайнозоя Казахстана начинается со среднего олигоцена. Начальный этап образования континентальных толщ с наземной фауной в этой области замечательно совладает с общим началом роста горных сооружений Средней Азии.
А.А. Борисяк подчеркивает, «что, начиная с олигоцена, фауны обеих стран (Азиатской части Союза и Монголии) имеют много общего, если не тождественны между собой».
В итоге сравнительного рассмотрения документов наземной жизни Монголии и нашей Средней Азии и Казахстана становится вероятным общее положение о большом периоде размыва, захватившем громадную площадь верхнемелового континента, уничтожившем широтную полосу местонахождений динозавров верхов мела и не допустившем аккумуляции континентальных эоценовых осадков. Этот эоценовый период размыва должен был быть связан не только с общими климатическими условиями, но с широко развитыми эпейрогеническими движениями. Для последних в нашем случае имеет значение не столько их вертикальный размах, сколько широкое распространение и синхронность. В настоящее время имеется мало данных о тектонических движениях в эоцене Средней Азии. Мы знаем о широком распространении палеогенового моря в районах морской трансгрессии, но история участков суши в эоцене совершенно темна. Некоторым указанием на проявление вулканизма могут служить покровы оливиновых базальтов в основании третичных континентальных толщ Иссык-кульской и других впадин, возможно принадлежащие эоцену. Напомним, что в Монголии мы встречаем проявления вулканизма в палеоцене (Гашато) и в верхах олигоцена (Хзанда-Гол).
В процессах образования мезокайнозойских костеносных континентальных толщ Средней Азии и Казахстана имеются и свои специфические черты, отличающие их от таких же процессов в Монголии. Признавая общность крупных этапов обеих великих областей, мы тем не менее должны отметить и качественную разницу областей захоронения фауны для будущего конкретного решения вопросов стратиграфии. Монголия в эпоху отложения костеносных свит мела и кайнозоя представляла собою область устойчивой депрессии внутри континента, с длительной аккумуляцией континентальных осадков. В этом внутриматериковом пространстве создались благоприятные условия для захоронения фауны, отразившей наземную, удаленную от морских берегов жизнь животного мира, ранее не известную.
Область распространения динозавровых местонахождений Средней Азии находилась в окраинной зоне Азиатского континента. Эта область имела условия, нужные для концентрации и захоронения фауны в береговой полосе мелового моря, в районах распространения крупных дельт. В этом отношении зона захоронения верхнемеловой фауны динозавров Средней Азии имела много сходства с крупнейшими динозавровыми местонахождениями Северной Америки и Европы. Полоса распространения динозавровых местонахождений имеет явственно широтное простирание, точнее простирание восток — северо-восток — запад — юго-запад. Параллельно этой полосе, далее к северу простирается полоса распространения местонахождений более поздних фаун млекопитающих — олигоценовой и миоценовой. Вряд ли здесь мы имеем дело со случайным совпадением. Нам представляется более вероятным предположение о закономерной миграции дельтовых поясов от гор на север в связи с общим прогрессирующим поднятием горных сооружений Средней Азии. Полоса распространения дельтовых фаций верхнего мела находилась несколько южнее остаточных аккумуляций великого эоценового размыва — современных динозавровых конгломератов. Далее к северу передвинулись в олигоцене и миоцене дельтовые фации индрикотериевых слоев и других более поздних фаун млекопитающих. Эта общая закономерность перемещения зон захоронения в мезокайнозое Средней Азии, разумеется, очерчена лишь в самых грубых чертах и, несомненно, имела местные отклонения. Однако намеченная картина имеет значение для прогнозов в поисках новых местонахождений и отражает постепенный рост среднеазиатских горных сооружений в древнетретичное время.
Для уточнения вопроса о граничных слоях мела и эоцена остается лишь провести сравнение с разрезами дельтовых поясов захоронения США и Европы. Возьмем наиболее известный разрез мезокайнозоя в горах Крейзи, штата Монтана, США. Здесь мы имеем весьма постепенный переход мощных континентальных отложений верхов мела в базальтные слои эоцена. Серию согласно залегающих меловых отложений венчает свита Ларами или цератопсовые слои — пески, глины и песчаники — мощностью около 400 м. На свиту Ларами совершенно согласно, с постепенным переходом слоев налегает формация Форт Юнион, сложенная сходными песчано-глинистыми отложениями и представляющая наиболее низкие горизонты эоцена. В самых нижних слоях нижнего песчано-глинистого отдела Форт Юнион находится фауна древнеэоценовых млекопитающих (Claenodon, Mioclaenus). Эти слои непосредственно подстилаются ларамийскими слоями с фауной беспозвоночных, характерных для цератопсовых слоев. Ниже горизонта с фауной беспозвоночных залегает свита глинистых пород с обильной эоценовой флорой, характерной для Форт Юнион. Ниже снова идут слои с моллюсками и крупными динозаврами — цератопсидами и траходонтами (Triceratops, Trachodon). Мы видим в этом разрезе отчетливое сближение фаун динозавров и млекопитающих, великая смена которых проходит при общности флоры и крайнем сходстве беспозвоночных, другими словами, без всяких резких изменений физико-географической обстановки.
Такой спокойный и весьма постепенный переход, знаменующий величайшую смену континентальных фаун, доказывает неразрывность общих процессов геологической истории на границе мезозоя и кайнозоя для некоторых областей древних континентов. В морских фаунах этот переход еще более незаметен.
Местонахождения среднеазиатских динозавров первой группы, подвергавшиеся перемыву в условиях близости береговой линии, окружены отложениями с морской фауной, как мелового, так и эоценового характера. Выше мы отмечали, что стратиграфическое положение динозаврового горизонта в сводном разрезе мезокайнозоя Приташкентского района является еще далеко неясным. Однако близость в разрезе морских отложений с фауной мелового типа является несомненной. По-видимому, для этой части Средней Азии общий размыв дельтовых осадков с местонахождениями динозавров мог наступить несколько раньше, что становится вполне понятным, если допустить длительное существование в этом районе крупной водной артерии. В то же время благодаря близости базиса эрозии — моря — перемыв не был настолько глубок, как в восточной части полосы динозавровых местонахождений, и мог сохранить часть пород, в свое время подстилавших коренные местонахождения динозавров. Таким образом, динозавровые местонахождения первой группы составляли краевую зону эоценовой суши и, вероятно, частично покрывались эоценовым морем.
Во всяком случае, все рассмотренные нами данные говорят о повышении возраста динозаврового горизонта Приташкентского района против принимавшегося до сих пор. Нужно думать, что если известняки с морской фауной дарбазинской свиты действительно залегают выше динозаврового горизонта, то определение их возраста как сенонского является сомнительным и вообще корреляция разрезов мела и палеогена Приташкентского района с классическими разрезами нуждается в уточнении. Если горизонты с меловой фауной окажутся залегающими под костеносными породами динозаврового горизонта, то они могут иметь любой, далекий от эоцена, возраст, попадая под уровень неглубокого размыва области местонахождений первой группы. На границе мела с эоценом и в континентальных, а тем более в морских слоях мы очень часто встречаемся с переплетением слоев, охарактеризованных фаунистическими группировками то мелового, то третичного облика. Сходные соотношения имеют место в Приташкентском районе и еще ждут подробного анализа.
В заключение остается сказать несколько слов о возможности находок континентальных фаун наземных позвоночных мела и эоцена в коренном залегании. Выше мы приближенно наметили схему распространения дельтовых фаций с остатками наземной фауны в виде полос, постепенно продвигавшихся на север и налегавших на морские осадки в пределах палеогеновой морской трансгрессии или же прямо на денудационную поверхность на субаэральвых участках суши. В первом приближении поиски местонахождений должны быть произведены по простиранию этих полос захоронения, сообразуясь с тектонической историей ближайших горных сооружений в соответствующее время.
Для меловых континентальных отложений, сохранившихся от размыва в больших депрессиях типа Ферганской (впадины, характерно, что их верхние свиты венчаются морскими осадками с морской фауной верхнего мела. По всей вероятности, эти морские осадки соответствуют континентальным, содержавшим фауну динозавров и подвергшимся размыву. Действительно, в северо-восточном направлении для Ферганской впадины отмечается переход морской верхней свиты мела в континентальную. Период захоронения фауны динозавров пришелся на верхи мела, поэтому в нижележащих свитах трудно рассчитывать на местонахождения динозавровой фауны. В условиях изменчивости типов осадков, как правило, не бывает близко следующих друг за другом крупных периодов захоронения. По всем этим соображениям мы должны искать коренные местонахождения динозавров в континентальных свитах самых верхов мела на востоке и северо-востоке от Ферганской впадины. Вследствие существовавшего о этой области великого эоце-нового размыва такие свиты могут быть обнаружены только в депрессиях, устойчивых с верхнемелового времени и выраженных в рельефе относительно меловой денудационной поверхности. Подобные условия могут быть обнаружены в восточном конце Илийской котловины (область Кульджи), также в Иссыккульской впадине, в последней, однако, в наиболее глубоких ее частях, покрытых водой озера.
Перспективы обнаружения местонахождений эоценовой континентальной фауны более темны, поскольку палеогеография эоценовой суши Средней Азии еще крайне неясна. Не считая погруженных под толщу более молодых осадков, продуктов крупного эоценового размыва, пока недоступных нашему исследованию на севере в больших впадинах Казахстана, мы можем обнаружить участки с эоценовой континентальной аккумуляцией в крупных древних впадинах, находившихся в пределах окраины эоценовой суши, поднятых и вскрытых позднейшими движениями. К таковым ориентировочно могут принадлежать Кульджинская котловина, впадина озера Зайсан и другие подобные образования.
Заключение
В итоге рассмотренного нами вопроса о возрасте и генезисе динозавровых местонахождений Средней Азии мы приходим к следующим основным выводам. Однако фактический материал, лежащий в основе наших сопоставлений, недостаточен, поэтому предлагаемые выводы следует рассматривать как предварительные соображения, намечающие и облегчающие путь дальнейшего изучения очень интересного и важного вопроса об истории суши в Средней Азии на границе мезозоя и кайнозоя.
1. Стратиграфическое значение остатков динозавров, ввиду плохой сохранности, не допускающей детального изучения, является условным.
2. Возраст динозавровой фауны для всех местонахождений должен быть в общем однозначным и приблизительно соответствует отрезку времени сенон — датский ярус (ларами), в среднем, верхам сенона,
3. Возраст местонахождений динозавров вследствие вторичного залегания остатков во всех без исключения местонахождениях безусловно моложе возраста самой фауны.
4. Для местонахождений западной части полосы распространения динозавровых конгломератов (первая группа — Приташкентский район и Кызыл-кумы) их возраст сравнительно близок к возрасту фауны; для местонахождений восточной части указанной полосы (вторая группа: местонахождения Туз-куль, Бозабы, Кок-Муйнак, Карой и т. д.) — более далек.
5. Местонахождения динозавров образовались в результате перемыва коренных дельтовых верхнемеловых фаций, простиравшихся в виде почти широтного пояса на северной окраине горных сооружений Средней Азии. Этот перемыв следует рассматривать как длительный процесс, обусловленный совокупностью физико-географических условий суши на переходе от мезозоя к кайнозою в Средней Азии.
6. Резкое выражение континентальной обстановки и интенсивность перемыва характерны для восточной области полосы динозавровых местонахождений. В западной части при близком расположении дельтовых осадков верхнего мела к базису эрозии перемыв не был глубоким и, начавшись ранее, быстро окончился.
7. Возраст процесса размыва предположительно определяется как лалеоцен — эоцен.
8. Процесс размыва определил отсутствие континентального эоцена а областях аккумуляции третичных осадков на древней денудационной поверхности мезокайнозойской суши.
9. Совокупностью седиментационных условий мезокайнозоя определяется отсутствие местонахождений эоценовой фауны млекопитающих на территории Средней Азии, включая Казахстан.
10. Местонахождения эоценовых млекопитающих могут быть найдены вне пределов области эоценовой суши, подвергшейся размыву и выносу продуктов осадконаколления и остатков наземных животных, или же в крупных древних депрессиях в пределах этой суши. Местонахождения динозавров в коренном залегании могут быть найдены в опущенных в эоценовое время участках верхнемелового дельтового пояса.
11. Возраст соответствующих слоев Приташкентского района (чанакская свита, дарбазинская свита и собственно динозавровый горизонт) представляется более молодым, чем это принимается в настоящее время. Во всяком случае динозавровый горизонт моложе сенона, а дарбазинская свита, если она залегает действительно над костеносными слоями, может принадлежать уже эоцену. Не исключена возможность эоценового возраста и самого динозаврового горизонта Приташкентского района.
Литература
1. Борисяк А.А. Обзор местонахождений третичных наземных млекопитающих. Союза ССР. Фрунзе, 1943.
2. Боркеман Б.А. Меловые отложения юго-востока Средней Азии. Ташкент, УзФАН, 1940.
3. Ефремов И.А. Динозавры в красноцветных толщах Средней Азии. Тр. Палеонт. инст. АН СССР, т. I, 1931. 4. Ефремов И.А. Два поля смерти минувших геологических эпох. Природа. № 7, 1933.
5. Минакова Н. Е. К стратиграфии меловых и третичных отложений Чулей. Ташкент, УзФАН, 1941.
6. Рябинин А.Н. Фауна позвоночных из верхнего мела Южного Казахстана. Tp. Центр. научн.-иссл. геол.-разв. инст. вып. 118.
7. Оsbогn H.F. Age of Mammals. New York, 1910.
8. Osborn H.P. Summary of Mongolian formations, faunas and geologic history
The New Conquest of Central Asia. New York, 1932.
Summary
In consequence of a consideration of the problem as to the age and origin of Dinosaur localities in Central Asia we come to the following conclusions. Reservation should be made, however, that the data laid at the basis of our correlations are quite insufficient. Therefore the conclusions arrived at should be considered as preliminary considerations marking and facilitating the way of further studies concerning the very interesting and important problem of the history of the continent in Central Asia at the junction of the Mesozoic and the Cenozoic.
1. The stratigraphic significance of the remains of dinosaurs is conditional owing to the bad preservation forbidding a detailed study.
2. The age of dinosaur fauna of all the localities must be about the same, belonging approximately to the Senonian-Danian (Laramie), most likely to the Upper Senonian.
3. The dinosaur localities are doubtlessly younger than the fauna itself, in consequence of a secondary embedding of the remains in all the localities without exception.
